BRAIN CHOLESTEROL PATHOLOGY IS THE CAUSE OF ALZHEIMERS DISEASE
Alexei R. Koudinova,CA and Natalia V. Koudinovaa,b
a Berezov Academic Laboratory, Russian Academy of Medical
Sciences, Timoshenko St., 38-27, Moscow, 121359 Russia;
b Weizmann Institute of Science, Department of Biological
Regulation, Rehovot, 76100 Israel.
First submitted: January 5, 2001, Published online: November, 2001
| COMPANION ARTICLE |
|
|
In accord with the above papers [ 6, 7 ] and despite of tolerating hypercholesterolemia [ 12 ] albino wistar rats fed a cholesterol diet demonstrate increased cholesterol and phospholipid synthesis in the hippocampus (Fig. 1, Table) and are characterized by Alzheimers-like amyloid (Figs. 1B, C, E; thus being a phenomenon secondary to brain cholesterol abnormality and possible aiming to modulate impaired neural cholesterol dynamics [ 11, 15 ], see below) and impaired long-term potentiation (LTP, Fig. 1F), a long-lasting plastic changes of synapses that underlie learning and memory [ 8, 11 ]. Moreover, impaired synaptic plasticity in rat ex-vivo hippocampal slices is caused by experimentally inhibited cholesterol synthesis [ 17 ] or increased cholesterol efflux [ 11 ]. The later condition in vivo may well be due to functionally, abnormally or experimentally increased rate of brain cholesterol synthesis (Fig. 1, Table) and associated turnover upregulation [ 18 ].
The experimental condition of increased cholesterol efflux is also characterized by paired helical filaments (PHF)-tau hyperphosphorylation in neurofibrillary tangles (NFT) and neurite degeneration [ 11 ], another key histochemical feature of AD. The observation of cholesterol-dependent tau phosphorylation in ex-vivo hippocampal slices [ 11 ] and in cultured neurons [ 19WEB+ ] as well as higher values of intracellular cholesterol in NFT-bearing neurons of AD cortex [ 20 ] the Alz-50 (an antibody that recognizes tau in NFTs) immunoreactivity in the brain of cholesterol-fed rabbits [ 7 ] suggest that NFT formation is a secondary phenomenon of abnormal neuronal cholesterol homeostasis [ LE1 ].
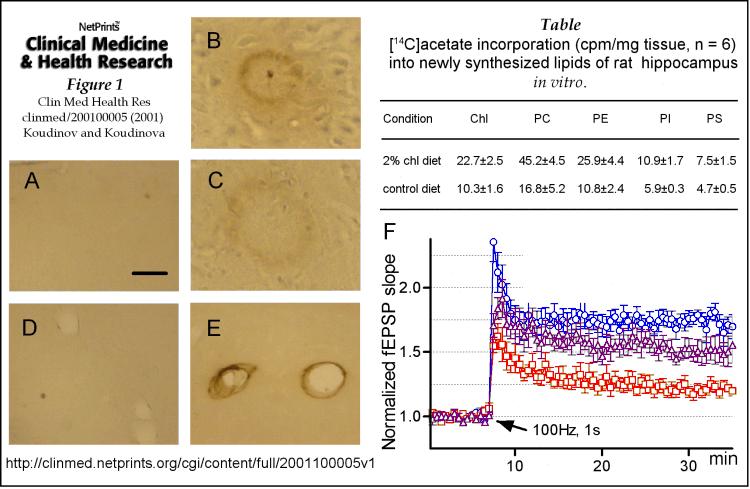
Consistent with previous data [ 8, 10, 12, 13, 14 ] by chronic modification of adult albino Wistar rat cholesterol status with a cholesterol diet we generated an animal model, characterized by significantly increased hippocampal synthesis of cholesterol and phospholipids, two major lipid components of the membranes of any living cell and of neurons, where nervous system activity is generated and propagated. We investigated cholesterol and different phospholipids syntheses in ex-vivo hippocampal slices of cholesterol-fed and control rats by incorporation of [14C]-acetate precursor into the newly synthesized lipid species, followed by lipid extraction, lipid separation by TLC, and radioactivity counting. This experimental protocol was previously employed and adapted for the ex-vivo brain slices [ 11 ]. In all cases the data were presented as lipid-incorporated radioactivity, mean ± SEM (n=6). As illustrated in the Table, cholesterol diet significantly upregulated hippocampal [and cortical (not shown)] cholesterol (chl) and all tested phospholipid syntheses (Mann-Whitney test, P<0.05, one tailed), indicating that dietary cholesterol causes brain cholesterol turnover upregulation in cholesterol-fed rats. PC, PE, PI and PS designate phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol and phosphatidylserine , respectively.
The following immunohistochemical analysis and electrophysiological experiments were performed as we detailed previously [ 11, 16 ]. Panels A-E show extracellular immunohistochemical staining of Ab with 4G8 (1:1000) monoclonal antibody [ 11 ] (anti-rodent/humanAb17-24) in brain sections of cholesterol fed rats (B, C, E). Alzheimer's-like plaque amyloid (B, C) and vascular Ab deposition (E) are illustrated in the hippocampus (E) and cortex (B, C). Panels A and D represent control rat cortical and hippocampal fields, respectively. Bar, 50 mM for panels A-C; and 30 mM for panels D, E.
Panel F demonstrates impairment of tetanus induced long term potentiation (LTP) in CA1 of ex-vivo hippocampal slices in cholesterol-fed rats (squares, n=8 slices) versus control animals (circles, n=12). LTP, a characteristic of synaptic plasticity [ 11, 16 ], was expressed as a normalized field excitatory postsynaptic potential (fEPSP) slope change versus time. Arrow indicates the time of tetanic stimuli train. Note that reversal of cholesterol diet to control diet for 5 weeks caused significant (P<0.05, one tailed) reversal of LTP impairment (triangles, n=5).
Presented data thus provide experimenal evidence that hippocampal cholesterol dynamics dysfunction causes Alzheimers major functional phenomenon of synaptic plasticity impairment and major pathological hallmark of amyloid deposition. Impairment of neurotransmission and synaptic plasticity in acute model of brain cholesterol pathology lacking Ab deposition (that we reported recently in The FASEB Journal [ 11 ] indicates that cholesterol (and not Ab) misregulation is a primary cause of synaptic dysfunction. Moreover, as lipid metabolism functional player, Ab likely modulates its biology in presented here chronic model of cholesterol-fed rat in order to recover impaired rat brain cholesterol homeostasis (see below).
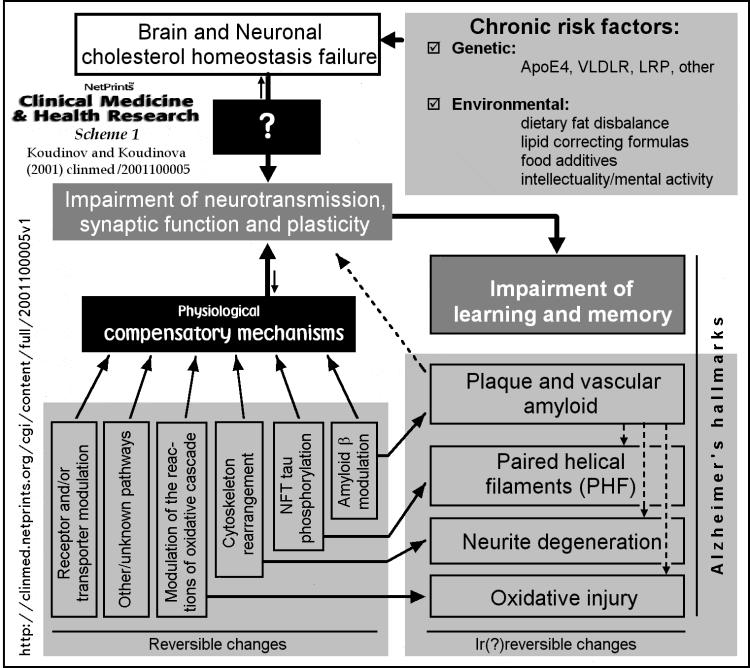
Conceivable, the break in any element of this harmonized system (caused by genetic defects of one of the associated proteins or by non-genetic [environmental, for example] factors) may result in abnormal homeostasis of cholesterol in the brain, impair fine tuning of neuronal functione and cause Alzheimers-like neurodegeneration features (Fig. 1 and Scheme 1) [ 6, 8, 11, 15, 17, 25, 26, 27WEB+ ]. Importantly, crosstalk of hepatic and neuronal cholesterol (see previously and [ 10, 12, 13, 14 ]) makes systemic cholesterol imperfection an important factor in developing brain cholesterol homeostasis failure. Several clinical and experimental reports favor relevance of the drawn possibility to the disease. Thus, first, AD is characterized by reduced cholesterol esterification [ 28, 29, 30 ] implying an abnormal CSF high density lipoprotein interconversion, critical for extracellular cholesterol trafficking and reverse cholesterol transport [ 23 ]. Second, AD is characterized by activation of the pathway for cholesterol removal out of the brain based on its oxidative conversion into 24S-hydroxycholesterol [ 18 ], indicating insufficiency and/or saturation of lipoprotein-mediated cholesterol disposal [ 23 ]. Third, cholesterol accumulates in senile plaques of AD patients and in plaque-like amyloid of aged mutated amyloid precursor protein transgenic mice [ 31 ], suggesting the interrelation of the repartitioning of cholesterol in the brain and amyloid deposition. Forth, human ApoE e4/e4 knock-in mice have markedly altered systemic and brain cholesterol metabolism [ 32 ], offering cholesterol trafficking attenuation as an explanation for the increased AD risk in ApoE e4/e4 subjects. Fifth, recent observation documented significant elevation of the LDL receptor related protein [ LE2 ] levels in AD frontal cortex [ 33 ] opening the possibility of upregulation or insufficiency of lipoprotein-receptor-mediated neuronal cholesterol redistribution in the disease [ 11, 18, 23 ]. Finally, related to AD Down syndrome (DS), a trisomy 21, is copying Alzheimers cholesterol esterification abnormality [ 34 ]. DS is also characterized by a specific pathway of the liver sterol regulatory element binding protein (SREBP) activation with sterol-independent maturation of SREBP-1, and by high circulating and tissue cholesterol levels in the fetuses [ 35 ], implying importance of developmental fetal cholesterol pathology for earlier (compared to AD) genesis of Alzheimers features in trisomy 21 subjects.
The biochemical relation of cholesterol and Ab, however, is bidirectional, and the modulation of neuronal cholesterol dynamics by Ab likely has important functional consequences.
Particularly, Ab modulates neuronal cholesterol esterification [ 37, 38 ], influx [ 11, 39 ] and efflux [ 23, 40 ] and thus possibly regulates neural cholesterol intracellular compartmentation and extracellular trafficking [ 21WEB+, 23, 40, 41 ] Ab also modulates neuronal physical property of membrane fluidity [ 42, 43 ] suggested to be important for cholesterol-dependent cell receptor machinery impairment (discussed in Ref. 11). Additionally, Ab increases lipid synthesis (specifically that of cholesterol and phospholipids) in PC12 and rat primary neuronal cell cultures, fetal brain, and in ex vivo hippocampal slices [ 11, 44, 45 ] in contrast to the peptide inhibitory (and cholesterol lowering [ 46 ]) effect, observed in human hepatic HepG2 and in HEK293 cells, in fetal rat liver and in neuronal tissue under the condition of potassium-evoked depolarization and under oxidative stress [ 30, 44, 45, 46 ]. The latter results highlight the importance of developmental, tissue and neuronal functional specificity of Ab-cholesterol biochemical relation, which may vary in different brain regions and be of special importance in determining Alzheimer's specific areas of neurodegeneration [ 30, 44, 45 ]. These data also suggest that Ab may serve a molecular messanger function and manage the crosstalk of hepatic, systemic and brain cholesterol, and thus maintain the tissue-specific coordinate regulation of cholesterol biosynthesis [ 10, 12, 13, 14 ]. Taken together, the above functional consideration and recent data on the importance of cholesterol compartmentation for Ab generation [ 2, 41 ] indicate feedback functional relation between cholesterol and Ab homeostasis, additionally supported by a dependency of amyloid precursor protein processing and Ab production on the site 2 processing of SREBP [ 47 ] (the major regulatory protein in cholesterol metabolism [ 21WEB+, 24, 35]) and associated inability of cells to upregulate the expression of several enzymes and proteins involved in cholesterol synthesis and turnover [ 47 ].
We believe that the fundamental pathophysiological
event in most common sporadic forms of AD is accurate brain cholesterol
homeostasis failure.
( See Scheme 1 )
FOR
YOUR CONVENIENCE:
|
THIS
ARTICLE IS BASED ON:
|
|
1. ( 4 web+ citations ) Marx J. Alzheimer's disease: bad for the heart, bad for the mind? Science. 294, 508-509 [ PubMed ] [ Full Text ]; Wolozin B. A fluid connection: Cholesterol and Ab. Proc Natl Acad Sci USA. 98, 5371-5373 (2001) [ PubMed ] [ Full Text ]; Simons M, Keller P, Dichgans J, Schulz JB. Cholesterol and Alzheimers disease: Is there a link? Neurology. 57, 1089-1093 (2001) [ Pub Med ] [ Full Text ]; Golde TE, Eckman CB. Cholesterol modulation as an emerging strategy for the treatment of Alzheimer's disease. Drug Discovery Today. 6, 1049-1055 (2001) [ PubMed ] [ Full Text ] [ Letter to the Editor ].
2. Puglielli L, Konopka G, Pack-Chung E, et al. Acyl-coenzyme A: cholesterol acyltransferase modulates the generation of the amyloid beta-peptide. Nature Cell Biol. 10, 905-912 (2001) [ PubMed ] [ Full text ] [ Letter to the Editor ].
3. Jick H, Zornberg GL, Jick SS, Seshadri S, Drachman, D.A. Statins and the risk of dementia. Lancet. 356, 1627-1631 (2000) [ PubMed ] [ Full Text ] [ Letter to the Editor ].
4. Wolozin B, Kellman W, Ruosseau P, Celesia GG, Siegel G. Decreased prevalence of Alzheimer disease associated with 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors. Arch Neurol. 57, 1439-1443 (2000) [ PubMed ] [ Full Text ] [ Letter to the Editor ].
5. Haley RW, Dietschy JM. Is there a connection between the concentration of cholesterol circulating in plasma and the rate of neuritic plaque formation in Alzheimer disease? Arch Neurol. 57, 1410-1412, (2000) [ PubMed ].
6. Refolo LM, Pappolla MA, Malester B, et al. Hypercholesterolemia accelerates the Alzheimers amyloid pathology in a transgenic mouse model. Neurobiol Dis. 7, 321-331 (2000) [ PubMed ] [ Full Text ].
7. Sparks DL, Scheff SW, Hunsaker JC 3rd, Liu H, Landers T, Gross DR. Induction of Alzheimer-like beta-amyloid immunoreactivity in the brains of rabbits with dietary cholesterol. Exp Neurol. 126, 88-94 (1994) [ PubMed ].
8. Senturk UK, Oner G. The effect of manganese-induced hypercholesterolemia on learning in rats. Biol Trace Elem Res. 51, 249-257 (1996) [ PubMed ].
9. Eckert GP, Kirsch C, Muller WE. Differential effects of lovastatin treatment on brain cholesterol levels in normal and ApoE-deficient mice. Neuroreport. 12, 883-887 (2001) [ PubMed ] [ Full Text ].
10. Edmond J, Korsak RA, Morrow JW, Torokboth G, Catlin DH. Dietary-cholesterol and the origin of cholesterol in the brain of developing rats. J Nutrition. 121, 1323-1330 (1991) [ PubMed ].
11. Koudinov AR, Koudinova NV. Essential role for cholesterol in synaptic plasticity and neuronal degeneration. FASEB J. 15, 1858-1860 (2001), published online June 27, 2001, 10.1096/fj.00-0815fje [ PubMed Citation ] [ Full Text ] [ Reprint Order ].
Cited above companion article is a prologue for this report. It discusses the role for cholesterol, Ab and tau in synaptic function and plasticity
12. Roach PD, Balasubramaniam S, Hirata F, et al. The low-density lipoprotein receptor and cholesterol synthesis are affected differently by dietary cholesterol in the rat. Biochim Biohys Acta. 1170, 165-172 (1993) [ PubMed ].
13. Morell P, Jurevics H. Origin of cholesterol in myelin. Neurochem Res. 21, 463-470 (1996) [ PubMed ].
14. Spady DK, Cuthbert JA. Regulation of hepatic sterol metabolism in the rat. Parallel regulation of activity and mRNA for 7 alpha-hydroxylase but not 3-hydroxy-3-methylglutaryl-coenzyme A reductase or low density lipoprotein receptor. J Biol Chem. 267, 5584-5591 (1992) [ PubMed ].
15. Sparks DL. Increased cholesterol in AD and cCAD cortex, and increased cholesterol content and free radical enzyme activity in rabbit brain due to dietary cholesterol. J Neuropath Exp Neurol. 55, 51 (1996).
16. Friedman LK, Koudinov AR. Unilateral GluR2(B) Hippocampal Knockdown: A Novel Partial Seizure Model in Young Rat. J Neurosci. 19, 9412-9425 (1999) [ PubMed ] [ Full Text ].
17. Matthies H, Schulz S, Hollt V, Krug M. Inhibition by compactin demonstrates a requirement of isoprenoid metabolism for long-term potentiation in rat hippocampal slices. Neurosci. 79, 341-346 (1997) [ PubMed ] [ Full Text ].
18. Papassotiropoulos A, Lutjohann D, Bagli M, et al. Plasma 24S-hydroxycholesterol: a peripheral indicator of neuronal degeneration and potential state marker for Alzheimers disease. Neuroreport. 11, 1959-1962 (2000) [ PubMed ].
19 ( 4 web+ citations ). Fan QW, Yu W, Senda T, Yanagisawa K, Michikawa M. Cholesterol-dependent modulation of tau phosphorylation in cultured neurons. J Neurochem. 76, 391-400 (2001) [ PubMed ] [ Full Text ]; Bi X, Zhou J, Sharman K, Song Z, Liu J, Lynch G. Induction of Alzheimers disease type pathologies in cultured hippocampal slices by cholesterol depletion. Soc Neurosci Abst. 27, 963.17 (2001) [ Abstract at ScholarOne.com ]; Sawamura N, Gong JS, Garver WS, Heidenreich RA, Ninomiya H, Ohno K, Yanagisawa K, Michikawa M. Site-specific phosphorylation of tau accompanied by activation of mitogen-activated protein kinase (mapk) in brains of Niemann-Pick type C mice. Soc Neurosci Abst. 27, 425.6 (2001) [ Abstract at ScholarOne.com ]; Bendiske J, Bahr BA. Positive lysosomal modulation reverses the PFH-tau accumulation, microtubule destabilization-transport failure-synaptic decay cascade Soc Neurosci Abst. 27, 425.15 (2001) [ Abstract at ScholarOne.com ].
20. Distl R, Meske V, Ohm TG. Tangle-bearing neurons contain more free cholesterol than adjacent tangle-free neurons. Acta Neuropathol (Berl). 101, 547-554 (2001) [ PubMed ].
21. ( 3 web+ citations ) Fielding CJ, Fielding P. Intracellular cholesterol transport. J Lipid Res. 38, 1503-1521 (1997) [ PubMed ]; Simons K, Ikonen E. How cells handle cholesterol. Science. 290, 1721-1726 (2000) [ PubMed ] [ Full Text ]; Dietschy JM, Turley SD. Cholesterol metabolism in the brain. Curr Opin Lipidol. 12, 105-112 (2001) [ PubMed ].
22. Poirier J, Baccichet A, Dea D, Gauthier S. Cholesterol synthesis and lipoprotein reuptake during synaptic remodelling in hippocampus in adult rats. Neurosci. 55, 81-90 (1993) [ PubMed ].
23. Koudinov AR, Berezov TT, Koudinova NV. The levels of soluble amyloid beta in different high density lipoprotein subfractions distinguish Alzheimer's and normal aging cerebrospinal fluid: implication for brain cholesterol pathology? Neurosci Lett. 314, 115-118 (2001) [ PubMed ] [ Full Text ] [ PDF ] [ Reprint Order ].
The above article presents experimental data and discusses how an impairment of extracellular lipoprotein-mediated trafficking of brain cholesterol may be linked to synaptic function, neural apolipoproteins and Alzheimers disease
24. Brown MS, Goldstein JL. The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell. 89, 331-340 (1997) [ PubMed ].
25. Crews FT, Camacho A, Philips MI. Cholinergic stimulation of hippocampal pyramidal cells is inhibited by increasing membrane cholesterol. Brain Res. 261, 155-158 (1983) [ PubMed ].
26. Tolbert LM, Lameh, J. Human muscarinic cholinergic receptor Hm1 internalizes via clathrin-coated vesicles. J Biol Chem. 271, 17335-17342 (1996) [ PubMed ].
27 ( 2 web+ citations ). Scanlon SM, Williams DC, Schloss P. Membrane cholesterol modulates serotonin transporter activity. Biochem. 40, 10507-10513 (2001) [ PubMed ] [ Full text ]; Mauch DH, Nagler K, Schumacher S, et al. CNS Synaptogenesis Promoted by Glia-Derived Cholesterol. Science 294, 1354-1357 (2001) [ PubMed ] [ Full text ] [ PubMed of accompanying commentary ] [ Accompanying commentary Full Text ] [ Letter to the Editor ].
28. Knebl J, DeFazio P, Clearfield MB, et al. Plasma lipids and cholesterol esterification in Alzheimer's disease. Mech. Aging Dev. 73, 69-77 (1994) [ PubMed ].
29. Demeester N, Castro G, Desrumaux C, et al. Characterization and functional studies of lipoproteins, lipid transfer proteins, and lecithin:cholesterol acyltransferase in CSF of normal individuals and patients with Alzheimers disease. J Lipid Res. 41, 963-974 (2000) [ PubMed ].
30. Koudinov AR, Berezov TT, Koudinova NV. Alzheimer's amyloid beta and lipid metabolism: a missing link? FASEB J. 12, 1097-1099 (1998) [ PubMed ] [ Full text ] [ Reprint Order ].
The above article summarises our earlier contribution to the development of the concept of Ab as an essential normal human apolipoprotein having certain functions in lipid metabolism
31. Mori T, Paris D, Town T, et al. Cholesterol accumulates in senile plaques of Alzheimer disease patients and in transgenic APPsw mice. J Neuropathol Exp Neurol. 60, 778-785 (2001) [ PubMed ].
32. Hamanaka H, Katoh-Fukui Y, Suzuki K, et al. Altered cholesterol metabolism in human apolipoprotein E4 knock-in mice. Human Mol Gen. 9, 353-361 (2000) [ PubMed ].
33. Qiu ZH, Strickland DK, Hyman BT, Rebeck GW. Elevation of LDL receptor-related protein levels via ligand interactions in Alzheimer disease and in vitro. J Neuropath Exp Neurol. 60, 430-440 (2001) [ PubMed ] [ Letter to the Editor ].
34. Lacko AG, Hayes JD, McConathy WJ, Lacko I, Redheendran R. Lecithin: cholesterol acyltransferase in Down's syndrome. Clin Chim Acta. 132, 133-141 (1983) [ PubMed ].
35. Diomede L, Salmona M, Albani D, et al. Alteration of SREBP activation in liver of Trisomy 21 fetuses. Biochem Biophys Res Commun. 260, 499-503 (1999) [ PubMed].
36. Refolo LM, Pappolla MA, LaFrancois J, et al. A cholesterol-lowering drug reduces b-amyloid pathology in a transgenic mouse model of Alzheimer's disease. Neurobiol Dis. 8, 890-899 (2001) [ PubMed ] [ Full Text ].
37. Koudinov AR, Koudinova NV, Berezov TT. Alzheimer's peptides Ab1-40 and Ab1-28 inhibit the plasma cholesterol esterification rate. Biochem Mol Biol Inter. 38, 747-752 (1996) [ PubMed ] [ Reprint Order ].
38. Liu Y, Peterson DA, Schubert D. Amyloid beta peptide alters intracellular vesicle trafficking and cholesterol homeostasis. Proc Natl Acad Sci USA. 95, 13266-13271 (1998) [ PubMed ] [ Full text ].
39. Igbavboa U, Avdulov NA, Chochina SV, Sun GY, Wood WG. Amyloid beta peptides and cholesterol dynamics. Neurosci Lett. S55, S25 (2000).
40. Michikawa M, Gong JS, Fan QW, Sawamura N, Yanagisawa K. A novel action of Alzheimer's amyloid beta-protein (Abeta): oligomeric Abeta promotes lipid release. J Neurosci. 21, 7226-7235 (2001) [ PubMed ] [ Full Text ].
41. Yamazaki T, Chang TY, Haass C. Accumulation and aggregation of amyloid beta-protein in late endosomes of Niemann-Pick Type C Cells. J Biol Chem. 276, 4454-4460 (2001) [ PubMed ] [ Full Text ].
42. Chochina SV, Avdulov NA, Igbavboa U, Cleary JP, O'Hare EO, Wood WG. Amyloid beta-peptide(1-40) increases neuronal membrane fluidity. Role of cholesterol and brain region. J Lipid Res. 42, 1292-1297 (2001) [ PubMed ] [ Full Text ].
43. Muller WE, Kirsch C, Eckert GP. Membrane-disordering effects of beta-amyloid peptides. Biochem Soc Trans. 29, 617-623 (2001) [ PubMed ].
44. Koudinova NV, Koudinov AR, Yavin E. Alzheimers Ab1-40 peptide modulates lipid synthesis in neuronal cultures and intact rat fetal brain under normoxic and oxidative stress conditions. Neurochem Res. 25, 653-660 (2000) [ PubMed ] [ Abstract ] [ .PDF Full Text ] [ Reprint order ].
45. Koudinova N, Koudinov AR, Berezov T, Yavin E. Alzheimer's amyloid beta modulates lipid synthesis in rat hippocampus. Soc Neurosci Abst. 25, 1859 (1999) [ Abstract ] [ Presentation Order ].
46. Walters CE, Austen BM. Intracellular cholesterol and b-amyloid as factors in Alzheimers disease. Neurobiol Aging. 19, 49S (1998).
47. Manni ME, Cescato R, Paganetti PA. Lack of b-amyloid production in M19 cells deficient in site 2 processing of the sterol regulatory element binding proteins. FEBS Lett. 427, 367-370 (1998) [ PubMed ] [ Full Text ].
48. ( 2 web+ citations ) Brion JP, Smith C, Couck AM, Gallo JM, Anderton BH. Developmental changes in tau phosphorylation: fetal tau is transiently phosphorylated in a manner similar to paired helical filament-tau characteristic of Alzheimer's disease. J Neurochem. 61, 2071-80 (1993) [ PubMed ]; Fujita S, Okawa Y, Ikeda Y, Ishiguro K. Stress-induced reversible hyperphosphorylation of tau in the mouse brain. Soc Neurosci Abst. 27, 425.9 (2001) [ Abstract at ScholarOne.com ].
49. Taubes G. The soft science of dietary fat. Science. 291, 2536-2545 (2001) [ PubMed ] [ Full Text ].
50. ( 2 web+ citations ) Kessler AR, Yehuda S. Learning-induced changes in brain membrane cholesterol and fluidity: implications for brain aging. Int J Neurosci. 28, 73-82 (1985) [ PubMed ]; Kessler AR, Kessler B, Yehuda S. In vivo modulation of brain cholesterol level and learning performance by a novel plant lipid: Indications for interactions between hippocampal-cortical cholesterol and learning. Life Sci. 38, 1185-1192 (1986) [ PubMed ].
51. Grant WB, Clarke RP, Hendrie HC, Hall KS, Gao S, Ogunniyi A. Incidence of Dementia and Alzheimer Disease in Nigeria and the United States. JAMA. 85, 2448-2449 (2001) [ PubMed ].
52. ( 2 web+ citations ) Kokmen E, Cha RH, Rocca WA. Time trends in the prevalence of dementia and Alzheimers disease in Rochester, Minnesota. In: Iqbal K, Swaab DF, Windlad B, Wisniewski HM, eds. Alzheimer's disease and related disorders: etiology, pathogenesis and therapeutics. Chichester: John Wiley and Sons, Ltd. 3-7 (1999); Kokmen E, Beard CM, O'Brien PC, Kurland LT. Epidemiology of dementia in Rochester, Minnesota. Mayo Clin Proc. 71, 275 (1996) [ PubMed ].
53. Sparks DL. Intraneuronal beta-amyloid immunoreactivity in the CNS. Neurobiol Aging. 17, 291-299 (1996) [ PubMed ].
54. Mesulam M-M. Neuroplasticity failure in Alzheimer's disease: bridging the gap between plaques and tangles. Neuron. 24, 521-529 (1999) [ PubMed] [ Full Text ].
55. ( 2 web+ citations ) Zervas M, Dobrenis K, Walkley SU. Neurons in Niemann-Pick disease type C accumulate gangliosides as well as unesterified cholesterol and undergo dendritic and axonal alterations. J Neuropath Exp Neurol. 60, 49-64 (2001) [ PubMed ]; Xie C, Burns DK, Turley SD, Dietschy JM. Cholesterol is sequestered in the brains of mice with Niemann-Pick type C disease but turnover is increased.J Neuropathol Exp Neurol. 59, 1106-17 (2001) [ PubMed ].
56. Naslund J, Haroutunian V, Mohs R, et al. Correlation between elevated levels of amyloid b-peptide in the brain and cognitive decline. JAMA. 283, 1571-1577 (2000) [ PubMed ].
57. Koudinov AR, Koudinova NV, Berezov TT. Is Alzheimer's amyloid plaque formation a condition for neuronal dysfunction? Neurobiol Aging. 21, S155 (2000) [ Abstract ] [ Reprint Order ].
58. Chen G, Chen KS, Knox J, et al. A learning deficit related to age and beta-amyloid plaques in a mouse model of Alzheimer's disease. Nature. 408, 975-979 (2000) [ PubMed ].
59. Knowles RB, Wyart C, Buldyrev SV, et al. Plaque-induced neurite abnormalities: implications for disruption of neural networks in Alzheimer's disease. Proc Natl Acad Sci USA. 96, 5274-5279 (1999) [ PubMed ].
60. Kuperstein F, Reiss N, Koudinova NV, Yavin E. Biphasic modulation of protein kinase C and enhanced cell toxicity by amyloid peptide and anoxia in neuronal cultures. J Neurochem. 76, 758-767 (2001) [ PubMed ] [ Reprint order ].
61. Koudinov A, Groner Y, Segal M. Cu/Zn-SOD transgenic mice are impaired in slow onset, long term potentiation. Neurosci Lett. 51, S23 (1998) [ Abstract ] [ Presentation Order ].
62. Auerbach JM, Segal M. Peroxide modulation of slow onset potentiation in rat hippocampus. J Neurosci. 17, 8695-8701 (1997) [ PubMed ].
63. Perry G, Nunomura A, Hirai K, Takeda A, Aliev G, Smith MA. Oxidative damage in Alzheimer's disease: the metabolic dimension. Int J Dev Neurosci. 18, 417-421 (2000) [ PubMed ].
64. Michikawa M, Yanagisawa K. Inhibition of cholesterol production but not of nonsterol isoprenoid products induces neuronal cell death. J Neurochem. 72, 2278-2285 (1999) [ PubMed ].
65. Zhuo M, Holtzman DM, Li Y, et al. Role of tissue plasminogen activator receptor LRP in hippocampal long-term potentiation. J Neurosci. 20, 542-549 (2000) [ PubMed ] [ Letter to the Editor ].
66. Champagne D, Rochford J, Poirier J. ApoE and LDL receptor knockout mice display acquisition deficits in the Morris water maze. Soc Neurosci Abstr. 26, 1546 (2000) [ Abstract at ScholarOne.com ].
67. O'Brien WT, Xu G, Tint GS, Salen G, Servatius RJ. Blocking cholesterol synthesis impairs acquisition of the classically conditioned eyeblink response. Integr Physiol Behav Sci. 35, 120-131 (2000) [ PubMed ].
68. Xu G, Servatius RJ, Shefer S, et al. Relationship between abnormal cholesterol synthesis and retarded learning in rats. Metabolism Clin. Exp. 47, 878-882 (1998) [ PubMed ].
69. Muldoon MF, Barger SD, Ryan CM, et al. Effects of lovastatin on cognitive function and psychological well-being. Am J Med. 108, 538-546 (2000) [ PubMed ].
70. Koudinov AR, Koudinova NV. Modulation of cholesterol metabolism initiates Alzheimers amyloid deposition and neuronal dysfunction. Soc Neurosci Abstr. 26, 497 (2000) [ Abstract ].
71. Koudinov AR, Koudinova NV. Neuronal cholesterol pathology is the cause of Alzheimer's disease. Soc Neurosci Abst. 27, 23.3 (2001) [ Abstract at ScholarOne.com ].
RELATED
LETTERS TO EDITOR ![]()
[ Authors
"eLetters to Editor" collection ]
LE1. Koudinov AR, Koudinova NV. Alzheimer's pathogenesis: tau and amyloid - a consensus or a challenge for a third party quest ? Br Med J. E.letter published online September 4, 2001 [ Read the letter ].
LE2. Koudinov AR, Koudinova NV. LRP: a cholesterol recruitment checkpoint for neuronal structure-functional plasticity? J Clin Invest. E.letter published online October 12, 2001 [ Read the letter ].
LE3. Koudinov AR, Koudinova NV. Cholesterol, synaptic function and Alzheimer's disease. Br Med J. E.letter published online October 16, 2001 [ Read the letter ].
LE4. Koudinov AR, Berezov TT, Koudinova NV. Beware: the link is not simple. Neurology E.letter published online October 16, 2001 [ Read the letter ].
LE5. Koudinov AR, Koudinova NV. Dementia, cholesterol and the soft science of dietary fat. Br Med J. E.letter published online July 27, 2001 [ Read the letter ].
LE6. Koudinov AR. Pioneering new era of biomedical science publishing. Br Med J. E.letter published online October 8, 2001 [ Read the letter ].
LE7. Koudinov AR, Koudinova NV. (2001) Looking forward for a historic issue, or: Is there anything besides amyloid? Br Med J. E.letter published online October 19, 2001 [ Read the letter ].
LE8. Koudinov AR, Koudinova NV. (2001) Cholesterol supply and synaptic plasticity. Submitted to Science on November 9, 2001 [ Read the letter ].
1S. Bodovitz S, Klein WL. Cholesterol modulates alpha-secretase cleavage of amyloid precursor protein. J Biol Chem. 271, 4436-4440 (1996) [ PubMed ].
2S. Racchi M, Baetta R, Salviett N, et al. Secretory processing of amyloid precursor protein is inhibited by increase in cellular cholesterol content. 322, 893-898 (1997) [ PubMed ].
3S. Simons M, Keller P, De Strooper B, Beyreuther K, Dotti CG, Simons K. Cholesterol depletion inhibits the generation of beta-amyloid in hippocampal neurons. Proc Natl Acad Sci USA. 95, 6460-6464 (1998) [ PubMed ].
4S. Howland DS, Trusko SP, Savage MJ, et al. (1998) Modulation of secreted beta-amyloid precursor protein and amyloid beta-peptide in brain by cholesterol. J Biol Chem. 273, 16576-16582 (1998) [ PubMed ].
5S. Mizuno T, Haass C, Michikawa M, Yanagisawa K. Cholesterol-dependent generation of a unique amyloid b-protein from apically missorted amyloid precursor protein in MDCK cells. Biochim Biophys Acta. 1373, 119-130 (1998) [ PubMed ].
6S. Frears ER, Stephens DJ, Walters CE, Davies H, Austen BM. The role of cholesterol in the biosynthesis of b-amyloid. Neuroreport. 10, 1699-1705 (1999) [ PubMed ].
7S. Gouras GK, Xu H, Gross RS, et al. Testosterone reduces neuronal secretion of Alzheimer's beta-amyloid peptides. Proc Natl Acad Sci USA. 97, 1202-1205 (2000) [ PubMed ].
8S. Galbete JL, Martin TR, Peressini E, Modena P, Bianchi R, Forloni G.Cholesterol decreases secretion of the secreted form of amyloid precursor protein by interfering with glycosilation in the protein secretory pathway. Biochem J. 348, 307-313 (2000) [ PubMed ].
9S. Smith CC, Hyatt PJ, Stanyer L, Betteridge DJ. Platelet secretion of beta-amyloid is increased in hypercholesterolaemia. Brain Res. 896, 161-164 (2001) [ PubMed ].
10S. Fassbender K, Simons M, Bergmann C. Simvastatin strongly reduces levels of Alzheimer's disease beta-amyloid peptides Abeta 42 and Abeta 40 in vitro and in vivo. Proc Natl Acad Sci USA. 98, 5856-5861 (2001) [ PubMed ].
11S. Kojro E, Gimpl G, Lammich S, Marz W, Fahrenholz
F. Low cholesterol stimulates the nonamyloidogenic pathway by its effect
on the alpha- secretase ADAM 10. Proc Natl Acad Sci USA. 98,
5815-5820 (2001) [ PubMed
].
|
Readers statistics:
|